Which Way Is Rna Read During Translation
Proteins are synthesized from mRNA templates past a procedure that has been highly conserved throughout evolution (reviewed in Chapter three). All mRNAs are read in the five´ to 3´ direction, and polypeptide chains are synthesized from the amino to the carboxy terminus. Each amino acid is specified past three bases (a codon) in the mRNA, according to a nearly universal genetic code. The basic mechanics of protein synthesis are too the same in all cells: Translation is carried out on ribosomes, with tRNAs serving as adaptors between the mRNA template and the amino acids being incorporated into protein. Protein synthesis thus involves interactions between three types of RNA molecules (mRNA templates, tRNAs, and rRNAs), equally well as various proteins that are required for translation.
Transfer RNAs
During translation, each of the 20 amino acids must be aligned with their corresponding codons on the mRNA template. All cells contain a variety of tRNAs that serve equally adaptors for this process. As might be expected, given their common office in protein synthesis, unlike tRNAs share similar overall structures. Notwithstanding, they likewise possess unique identifying sequences that let the correct amino acid to be attached and aligned with the appropriate codon in mRNA.
Transfer RNAs are approximately 70 to 80 nucleotides long and have characteristic cloverleaf structures that consequence from complementary base pairing between different regions of the molecule (Effigy 7.1). X-ray crystallography studies have further shown that all tRNAs fold into like meaty L shapes, which are likely required for the tRNAs to fit onto ribosomes during the translation process. The adaptor function of the tRNAs involves 2 separated regions of the molecule. All tRNAs have the sequence CCA at their 3´ terminus, and amino acids are covalently attached to the ribose of the final adenosine. The mRNA template is and then recognized past the anticodon loop, located at the other end of the folded tRNA, which binds to the appropriate codon by complementary base pairing.
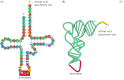
Effigy seven.i
Structure of tRNAs. The structure of yeast phenylalanyl tRNA is illustrated in open "cloverleaf" grade (A) to evidence complementary base of operations pairing. Modified bases are indicated as mG, methylguanosine; mC, methylcytosine; DHU, dihydrouridine; (more than...)
The incorporation of the correctly encoded amino acids into proteins depends on the attachment of each amino acid to an advisable tRNA, equally well equally on the specificity of codon-anticodon base pairing. The attachment of amino acids to specific tRNAs is mediated by a group of enzymes called aminoacyl tRNA synthetases, which were discovered by Paul Zamecnik and Mahlon Hoagland in 1957. Each of these enzymes recognizes a single amino acrid, as well as the correct tRNA (or tRNAs) to which that amino acid should be attached. The reaction proceeds in ii steps (Figure 7.2). First, the amino acid is activated past reaction with ATP to form an aminoacyl AMP synthetase intermediate. The activated amino acid is then joined to the 3´ terminus of the tRNA. The aminoacyl tRNA synthetases must be highly selective enzymes that recognize both individual amino acids and specific base sequences that place the correct acceptor tRNAs. In some cases, the loftier allegiance of amino acid recognition results in office from a proofreading function by which incorrect aminoacyl AMPs are hydrolyzed rather than beingness joined to tRNA during the second stride of the reaction. Recognition of the correct tRNA by the aminoacyl tRNA synthetase is besides highly selective; the synthetase recognizes specific nucleotide sequences (in most cases including the anticodon) that uniquely identify each species of tRNA.
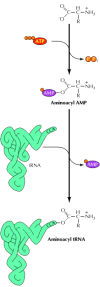
Figure 7.ii
Attachment of amino acids to tRNAs. In the first reaction step, the amino acid is joined to AMP, forming an aminoacyl AMP intermediate. In the second step, the amino acid is transferred to the 3´ CCA terminus of the acceptor tRNA and AMP is released. (more than...)
After being fastened to tRNA, an amino acrid is aligned on the mRNA template by complementary base pairing between the mRNA codon and the anticodon of the tRNA. Codon-anticodon base pairing is somewhat less stringent than the standard A-U and G-C base pairing discussed in preceding chapters. The significance of this unusual base pairing in codon-anticodon recognition relates to the redundancy of the genetic code. Of the 64 possible codons, three are stop codons that betoken the termination of translation; the other 61 encode amino acids (see Table iii.1). Thus, nearly of the amino acids are specified past more than i codon. In part, this back-up results from the attachment of many amino acids to more than 1 species of tRNA. E. coli, for case, comprise almost xl unlike tRNAs that serve as acceptors for the 20 different amino acids. In addition, some tRNAs are able to recognize more than than i codon in mRNA, as a result of nonstandard base of operations pairing (called wobble) betwixt the tRNA anticodon and the third position of some complementary codons (Effigy 7.3). Relaxed base of operations pairing at this position results partly from the formation of G-U base pairs and partly from the modification of guanosine to inosine in the anticodons of several tRNAs during processing (see Figure 6.38). Inosine can base-pair with either C, U, or A in the third position, so its inclusion in the anticodon allows a single tRNA to recognize iii unlike codons in mRNA templates.
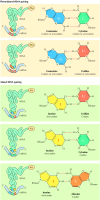
Effigy vii.3
Nonstandard codon-anticodon base of operations pairing. Base pairing at the third codon position is relaxed, assuasive Thousand to pair with U, and inosine (I) in the anticodon to pair with U, C, or A. Two examples of aberrant base of operations pairing, allowing phenylalanyl (Phe) tRNA (more...)
The Ribosome
Ribosomes are the sites of protein synthesis in both prokaryotic and eukaryotic cells. First characterized as particles detected past ultracentrifugation of cell lysates, ribosomes are usually designated according to their rates of sedimentation: 70S for bacterial ribosomes and 80S for the somewhat larger ribosomes of eukaryotic cells. Both prokaryotic and eukaryotic ribosomes are composed of two distinct subunits, each containing characteristic proteins and rRNAs. The fact that cells typically contain many ribosomes reflects the fundamental importance of protein synthesis in prison cell metabolism. E. coli, for instance, contain about 20,000 ribosomes, which account for approximately 25% of the dry weight of the jail cell, and rapidly growing mammalian cells comprise about 10 million ribosomes.
The full general structures of prokaryotic and eukaryotic ribosomes are like, although they differ in some details (Figure 7.four). The small subunit (designated 30S) of E. coli ribosomes consists of the 16S rRNA and 21 proteins; the large subunit (50S) is equanimous of the 23S and 5S rRNAs and 34 proteins. Each ribosome contains one re-create of the rRNAs and 1 copy of each of the ribosomal proteins, with i exception: One protein of the 50S subunit is nowadays in four copies. The subunits of eukaryotic ribosomes are larger and contain more proteins than their prokaryotic counterparts accept. The small-scale subunit (40S) of eukaryotic ribosomes is composed of the 18S rRNA and approximately 30 proteins; the large subunit (60S) contains the 28S, 5.8S, and 5S rRNAs and most 45 proteins.

Effigy 7.4
Ribosome structure. (A) Electron micrograph of East. coli 50S ribosomal subunits. (B–C) High resolution 10-ray crystal structures of 30S (B) and 50S (C) ribosomal subunits. (D) Model of ribosome structure. (E) Components of prokaryotic and eukaryotic (more than...)
A noteworthy feature of ribosomes is that they tin can exist formed in vitro by self-assembly of their RNA and poly peptide constituents. As first described in 1968 by Masayasu Nomura, purified ribosomal proteins and rRNAs tin be mixed together and, under appropriate conditions, will reform a functional ribosome. Although ribosome associates in vivo (specially in eukaryotic cells) is considerably more than complicated, the ability of ribosomes to self-gather in vitro has provided an important experimental tool, allowing analysis of the roles of private proteins and rRNAs.
Like tRNAs, rRNAs course characteristic secondary structures by complementary base of operations pairing (Figure 7.five). In clan with ribosomal proteins the rRNAs fold further, into distinct three-dimensional structures. Initially, rRNAs were thought to play a structural role, providing a scaffold upon which ribosomal proteins assemble. Nonetheless, with the discovery of the catalytic activity of other RNA molecules (e.g., RNase P and the self-splicing introns discussed in Affiliate 6), the possible catalytic role of rRNA became widely considered. Consequent with this hypothesis, rRNAs were establish to exist absolutely required for the in vitro assembly of functional ribosomes. On the other paw, the omission of many ribosomal proteins resulted in a decrease, but not a consummate loss, of ribosome activity.

Figure 7.5
The structure of 16S rRNA. Complementary base pairing results in the formation of a distinct secondary structure. (From S. Stern, T. Powers, L.-I. Changchien and H. F. Noller, 1989. Science 244: 783.)
Direct evidence for the catalytic action of rRNA beginning came from experiments of Harry Noller and his colleagues in 1992. These investigators demonstrated that the big ribosomal subunit is able to catalyze the formation of peptide bonds (the peptidyl transferase reaction) even after approximately 95% of the ribosomal proteins have been removed by standard protein extraction procedures. In contrast, handling with RNase completely abolishes peptide bond formation, providing strong support for the hypothesis that the formation of a peptide bail is an RNA-catalyzed reaction. Further studies have confirmed and extended these results by demonstrating that the peptidyl transferase reaction tin can exist catalyzed past synthetic fragments of 23S rRNA in the total absenteeism of any ribosomal protein. Thus, the central reaction of protein synthesis is catalyzed past ribosomal RNA. Rather than being the primary catalytic constituents of ribosomes, ribosomal proteins are now idea to facilitate proper folding of the rRNA and to enhance ribosome function by properly positioning the tRNAs.
The direct interest of rRNA in the peptidyl transferase reaction has of import evolutionary implications. RNAs are idea to take been the first self-replicating macromolecules (run across Chapter i). This notion is strongly supported by the fact that ribozymes, such as RNase P and cocky-splicing introns, can catalyze reactions that involve RNA substrates. The role of rRNA in the germination of peptide bonds extends the catalytic activities of RNA across self-replication to direct involvement in poly peptide synthesis. Additional studies betoken that the Tetrahymena rRNA ribozyme tin catalyze the attachment of amino acids to RNA, lending credence to the possibility that the original aminoacyl tRNA synthetases were RNAs rather than proteins. The ability of RNA molecules to catalyze the reactions required for poly peptide synthesis as well as for cocky-replication may provide an important link for understanding the early evolution of cells.
The Organization of mRNAs and the Initiation of Translation
Although the mechanisms of poly peptide synthesis in prokaryotic and eukaryotic cells are like, there are also differences, peculiarly in the signals that make up one's mind the positions at which synthesis of a polypeptide chain is initiated on an mRNA template (Figure vii.six). Translation does not simply begin at the five´ finish of the mRNA; information technology starts at specific initiation sites. The five´ final portions of both prokaryotic and eukaryotic mRNAs are therefore noncoding sequences, referred to as 5´ untranslated regions. Eukaryotic mRNAs usually encode only a single polypeptide chain, simply many prokaryotic mRNAs encode multiple polypeptides that are synthesized independently from distinct initiation sites. For case, the East. coli lac operon consists of three genes that are translated from the aforementioned mRNA (see Figure 6.8). Messenger RNAs that encode multiple polypeptides are chosen polycistronic, whereas monocistronic mRNAs encode a unmarried polypeptide chain. Finally, both prokaryotic and eukaryotic mRNAs end in noncoding three´ untranslated regions.
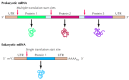
Figure seven.half-dozen
Prokaryotic and eukaryotic mRNAs. Both prokaryotic and eukaryotic mRNAs comprise untranslated regions (UTRs) at their v´ and 3´ ends. Eukaryotic mRNAs also contain five´ seven-methylguanosine (m7Yard) caps and 3´ poly-A tails. Prokaryotic (more...)
In both prokaryotic and eukaryotic cells, translation always initiates with the amino acid methionine, usually encoded past AUG. Culling initiation codons, such as GUG, are used occasionally in bacteria, but when they occur at the get-go of a polypeptide chain, these codons straight the incorporation of methionine rather than of the amino acid they normally encode (GUG normally encodes valine). In virtually bacteria, protein synthesis is initiated with a modified methionine balance (Due north-formylmethionine), whereas unmodified methionines initiate protein synthesis in eukaryotes (except in mitochondria and chloroplasts, whose ribosomes resemble those of leaner).
The signals that identify initiation codons are dissimilar in prokaryotic and eukaryotic cells, consistent with the distinct functions of polycistronic and monocistronic mRNAs (Figure 7.7). Initiation codons in bacterial mRNAs are preceded by a specific sequence (chosen a Shine-Delgarno sequence, after its discoverers) that aligns the mRNA on the ribosome for translation by base-pairing with a complementary sequence near the 3´ terminus of 16S rRNA. This base-pairing interaction enables bacterial ribosomes to initiate translation non only at the 5´ cease of an mRNA only too at the internal initiation sites of polycistronic messages. In contrast, ribosomes recognize most eukaryotic mRNAs by binding to the 7-methylguanosine cap at their 5´ terminus (see Figure 6.39). The ribosomes then scan downstream of the v´ cap until they run across an AUG initiation codon. Sequences that environment AUGs affect the efficiency of initiation, and so in many cases the first AUG in the mRNA is bypassed and translation initiates at an AUG farther downstream. However, eukaryotic mRNAs have no sequence equivalent to the Smoothen-Delgarno sequence of prokaryotic mRNAs. Translation of eukaryotic mRNAs is instead initiated at a site determined past scanning from the 5´ terminus, consistent with their functions equally monocistronic messages that encode only unmarried polypeptides.

Effigy 7.seven
Signals for translation initiation. Initiation sites in prokaryotic mRNAs are characterized by a Smooth-Delgarno sequence that precedes the AUG initiation codon. Base of operations pairing betwixt the Shine-Delgarno sequence and a complementary sequence near the 3´ (more than...)
The Process of Translation
Translation is by and large divided into 3 stages: initiation, elongation, and termination (Figure 7.eight). In both prokaryotes and eukaryotes the offset step of the initiation stage is the binding of a specific initiator methionyl tRNA and the mRNA to the pocket-size ribosomal subunit. The large ribosomal subunit then joins the complex, forming a functional ribosome on which elongation of the polypeptide chain proceeds. A number of specific nonribosomal proteins are also required for the various stages of the translation process (Table 7.i).
The outset translation step in leaner is the bounden of three initiation factors (IF-1, IF-2, and IF-3) to the 30S ribosomal subunit (Effigy seven.9). The mRNA and initiator N-formylmethionyl tRNA and so join the complex, with IF-2 (which is leap to GTP) specifically recognizing the initiator tRNA. IF-3 is and so released, allowing a 50S ribosomal subunit to associate with the circuitous. This association triggers the hydrolysis of GTP bound to IF-2, which leads to the release of IF-1 and IF-2 (bound to Gross domestic product). The effect is the formation of a 70S initiation complex (with mRNA and initiator tRNA leap to the ribosome) that is prepare to brainstorm peptide bond formation during the elongation stage of translation.
Effigy vii.9
Initiation of translation in bacteria. Three initiation factors (IF-one, IF-two, and IF-3) first bind to the 30S ribosomal subunit. This step is followed past bounden of the mRNA and the initiator N-formylmethionyl (fMet) tRNA, which is recognized by IF-2 bound (more...)
Initiation in eukaryotes is more complicated and requires at least ten proteins (each consisting of multiple polypeptide chains), which are designated eIFs (due eastukaryotic initiation factors; see Table seven.i). The factors eIF-i, eIF-1A, and eIF-3 bind to the 40S ribosomal subunit, and eIF-2 (in a circuitous with GTP) associates with the initiator methionyl tRNA (Effigy vii.x). The mRNA is recognized and brought to the ribosome by the eIF-4 grouping of factors. The 5´ cap of the mRNA is recognized by eIF-4E. Another factor, eIF-4G, binds to both eIF-4E and to a poly peptide (poly-A binding poly peptide or PABP) associated with the poly-A tail at the 3' finish of the mRNA. Eukaryotic initiation factors thus recognize both the 5' and iii' ends of mRNAs, accounting for the stimulatory effect of polyadenylation on translation. The initiation factors eIF-4E and eIF-4G, in association with eIF-4A and eIF-4B, then bring the mRNA to the 40S ribosomal subunit, with eIF-4G interacting with eIF-three. The 40S ribosomal subunit, in association with the leap methionyl tRNA and eIFs, then scans the mRNA to place the AUG initiation codon. When the AUG codon is reached, eIF-5 triggers the hydrolysis of GTP bound to eIF-2. Initiation factors (including eIF-2 spring to GDP) are and then released, and a 60S subunit binds to the 40S subunit to grade the 80S initiation circuitous of eukaryotic cells.

Figure vii.10
Initiation of translation in eukaryotic cells. Initiation factors eIF-3, eIF-1, and eIF-1A bind to the 40S ribosomal subunit. The initiator methionyl tRNA is brought to the ribosome by eIF-ii (complexed to GTP), and the mRNA by eIF-4E (which binds to the (more...)
After the initiation complex has formed, translation proceeds by elongation of the polypeptide chain. The machinery of elongation in prokaryotic and eukaryotic cells is very similar (Effigy 7.11). The ribosome has iii sites for tRNA binding, designated the P (peptidyl), A (aminoacyl), and Eastward (exit) sites. The initiator methionyl tRNA is bound at the P site. The start step in elongation is the binding of the next aminoacyl tRNA to the A site past pairing with the second codon of the mRNA. The aminoacyl tRNA is escorted to the ribosome by an elongation cistron (EF-Tu in prokaryotes, eEF-1α in eukaryotes), which is complexed to GTP. The GTP is hydrolyzed to Gdp as the correct aminoacyl tRNA is inserted into the A site of the ribosome and the elongation factor bound to Gross domestic product is released. The requirement for hydrolysis of GTP earlier EF-Tu or eEF-1α is released from the ribosome is the rate-limiting step in elongation and provides a time interval during which an incorrect aminoacyl tRNA, which would demark less strongly to the mRNA codon, can dissociate from the ribosome rather than being used for poly peptide synthesis. Thus, the expenditure of a loftier-energy GTP at this step is an important contribution to accurate poly peptide synthesis; it allows time for proofreading of the codon-anticodon pairing before the peptide bond forms.
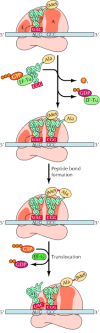
Figure vii.11
Elongation stage of translation. The ribosome has three tRNA-binding sites, designated P (peptidyl), A (aminoacyl), and Due east (get out). The initiating N-formylmethionyl tRNA is positioned in the P site, leaving an empty A site. The second aminoacyl tRNA (e.g., (more than...)
Once EF-Tu (or eEF-1α) has left the ribosome, a peptide bond can be formed betwixt the initiator methionyl tRNA at the P site and the second aminoacyl tRNA at the A site. This reaction is catalyzed past the large ribosomal subunit, with the rRNA playing a disquisitional function (as already discussed). The issue is the transfer of methionine to the aminoacyl tRNA at the A site of the ribosome, forming a peptidyl tRNA at this position and leaving the uncharged initiator tRNA at the P site. The adjacent step in elongation is translocation, which requires some other elongation factor (EF-K in prokaryotes, eEF-ii in eukaryotes) and is once more coupled to GTP hydrolysis. During translocation, the ribosome moves iii nucleotides along the mRNA, positioning the side by side codon in an empty A site. This step translocates the peptidyl tRNA from the A site to the P site, and the uncharged tRNA from the P site to the Eastward site. The ribosome is then left with a peptidyl tRNA bound at the P site, and an empty A site. The binding of a new aminoacyl tRNA to the A site so induces the release of the uncharged tRNA from the East site, leaving the ribosome prepare for insertion of the next amino acid in the growing polypeptide chain.
Equally elongation continues, the EF-Tu (or eEF-1α) that is released from the ribosome bound to Gross domestic product must be reconverted to its GTP class (Figure 7.12). This conversion requires a third elongation factor, EF-Ts (eEF-1βγ in eukaryotes), which binds to the EF-Tu/Gdp circuitous and promotes the exchange of bound GDP for GTP. This exchange results in the regeneration of EF-Tu/GTP, which is now set up to escort a new aminoacyl tRNA to the A site of the ribosome, offset a new cycle of elongation. The regulation of EF-Tu past GTP binding and hydrolysis illustrates a common means of the regulation of protein activities. As will exist discussed in afterwards chapters, like mechanisms control the activities of a wide diversity of proteins involved in the regulation of jail cell growth and differentiation, every bit well as in protein ship and secretion.
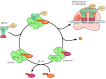
Figure 7.12
Regeneration of EF-Tu/GTP. EF-Tu complexed to GTP escorts the aminoacyl tRNA to the ribosome. The bound GTP is hydrolyzed every bit the correct tRNA is inserted, and so EF-Tu complexed to GDP is released. The EF-Tu/GDP complex is inactive and unable to demark another (more than...)
Elongation of the polypeptide chain continues until a stop codon (UAA, UAG, or UGA) is translocated into the A site of the ribosome. Cells practice not contain tRNAs with anticodons complementary to these termination signals; instead, they have release factors that recognize the signals and terminate protein synthesis (Figure 7.13). Prokaryotic cells comprise 2 release factors that recognize termination codons: RF-one recognizes UAA or UAG, and RF-two recognizes UAA or UGA (run into Table 7.one). In eukaryotic cells a unmarried release cistron (eRF-1) recognizes all iii termination codons. Both prokaryotic and eukaryotic cells also contain release factors (RF-3 and eRF-3, respectively) that practice not recognize specific termination codons just deed together with RF-1 (or eRF-one) and RF-2. The release factors bind to a termination codon at the A site and stimulate hydrolysis of the bond between the tRNA and the polypeptide chain at the P site, resulting in release of the completed polypeptide from the ribosome. The tRNA is so released, and the ribosomal subunits and the mRNA template dissociate.
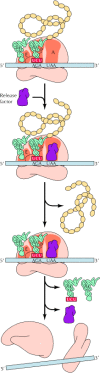
Figure 7.xiii
Termination of translation. A termination codon (east.thou., UAA) at the A site is recognized by a release factor rather than past a tRNA. The effect is the release of the completed polypeptide concatenation, followed past the dissociation of tRNA and mRNA from the ribosome. (more...)
Messenger RNAs can be translated simultaneously by several ribosomes in both prokaryotic and eukaryotic cells. One time one ribosome has moved away from the initiation site, another can bind to the mRNA and begin synthesis of a new polypeptide chain. Thus, mRNAs are usually translated by a series of ribosomes, spaced at intervals of nearly 100 to 200 nucleotides (Figure 7.fourteen). The group of ribosomes bound to an mRNA molecule is called a polyribosome, or polysome. Each ribosome within the group functions independently to synthesize a split polypeptide chain.

Effigy seven.xiv
Polysomes. Messenger RNAs are translated by a serial of multiple ribosomes (a polysome). (A) Electron micrograph of a eukaryotic polysome. (B) Schematic of a generalized poly-some. Note that the ribosomes closer to the three´ end of the mRNA take (more...)
Regulation of Translation
Although transcription is the primary level at which gene expression is controlled, the translation of mRNAs is likewise regulated in both prokaryotic and eukaryotic cells. One mechanism of translational regulation is the binding of repressor proteins, which block translation, to specific mRNA sequences. The best understood case of this mechanism in eukaryotic cells is regulation of the synthesis of ferritin, a poly peptide that stores iron inside the prison cell. The translation of ferritin mRNA is regulated by the supply of iron: More ferritin is synthesized if iron is abundant (Figure 7.15). This regulation is mediated by a protein which (in the absence of iron) binds to a sequence (the iron response element, or IRE) in the 5´ untranslated region of ferritin mRNA, blocking its translation. In the presence of iron, the repressor no longer binds to the IRE and ferritin translation is able to keep.

Figure 7.fifteen
Translational regulation of ferritin. The mRNA contains an iron response element (IRE) near its 5´ cap. In the presence of adequate supplies of fe, translation of the mRNA gain ordinarily. If iron is scarce, however, a protein (called the (more than...)
Information technology is interesting to note that the regulation of translation of ferritin mRNA past iron is like to the regulation of transferrin receptor mRNA stability, which was discussed in the previous affiliate (run across Figure 6.48). Namely, the stability of transferrin receptor mRNA is regulated by poly peptide binding to an IRE in its three´ untranslated region. The same protein binds to the IREs of both ferritin and transferrin receptor mRNAs. However, the consequences of protein binding to the two IREs are quite different. Protein spring to the transferrin receptor IRE protects the mRNA from degradation rather than inhibiting its translation. These distinct furnishings presumably consequence from the different locations of the IRE in the two mRNAs. To part as a repressor-binding site, the IRE must be located within 70 nucleotides of the 5´ cap of ferritin mRNA, suggesting that protein binding to the IRE blocks translation past interfering with cap recognition and binding of the 40S ribosomal subunit. Rather than inhibiting translation, protein binding to the same sequence in the 3´ untranslated region of transferrin receptor mRNA protects the mRNA from nuclease degradation. Binding of the same regulatory protein to unlike sites on mRNA molecules can thus have singled-out effects on gene expression, in one instance inhibiting translation and in the other stabilizing the mRNA to increase protein synthesis.
Another mechanism of translational regulation in eukaryotic cells, resulting in global effects on overall translational activity rather than on the translation of specific mRNAs, involves modulation of the activity of initiation factors, especially eIF-2. As already discussed, eIF-2 (complexed with GTP) binds to the initiator methionyl tRNA, bringing it to the ribosome. The subsequent release of eIF-2 is accompanied by GTP hydrolysis, leaving eIF-2 every bit an inactive GDP complex. To participate in another cycle of initiation, the eIF-2/GTP circuitous must be regenerated by the substitution of leap GDP for GTP. This exchange is mediated by another factor, eIF-2B. The control of eIF-2 activeness by GTP binding and hydrolysis is thus similar to that of EF-Tu (see Effigy 7.12). All the same, the regulation of eIF-ii provides a critical command betoken in a diversity of eukaryotic cells. In detail, eIF-2 tin can be phosphorylated by regulatory protein kinases. This phosphorylation blocks the exchange of bound GDP for GTP, thereby inhibiting initiation of translation. One blazon of cell in which such phosphorylation occurs is the reticulocyte, which is devoted to the synthesis of hemoglobin (Effigy 7.16). The translation of globin mRNA is controlled past the availability of heme: The mRNA is translated just if acceptable heme is available to grade functional hemoglobin molecules. In the absence of heme, a protein kinase that phosphorylates eIF-2 is activated, and farther translation is inhibited. Similar mechanisms have been found to control protein synthesis in other cell types, particularly virus-infected cells in which viral protein synthesis is inhibited by interferon.
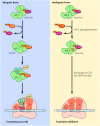
Figure seven.16
Regulation of translation by phosphorylation of eIF-2. Translation in reticulocytes (which is devoted to synthesis of hemoglobin) is controlled past the supply of heme, which regulates the activity of eIF-2. The agile form of eIF-ii (complexed with GTP) (more...)
Other studies take implicated eIF-4E, which binds to the 5´ cap of mRNAs, as a translational regulatory protein. For example, the hormone insulin stimulates protein synthesis in adipocytes and muscle cells. This effect of insulin is mediated, at least in office, past phosphorylation of proteins associated with eIF-4E, resulting in stimulation of eIF-4E action and increased rates of translational initiation.
Translational regulation is peculiarly of import during early development. As discussed in Chapter 6, a variety of mRNAs are stored in oocytes in an untranslated form; the translation of these stored mRNAs is activated at fertilization or later on stages of development. One mechanism of such translational regulation is the controlled polyadenylation of oocyte mRNAs. Many untranslated mRNAs are stored in oocytes with brusk poly-A tails (approximately 20 nucleotides). These stored mRNAs are afterwards recruited for translation at the appropriate stage of development by the lengthening of their poly-A tails to several hundred nucleotides. In improver, the translation of some mRNAs during development appears to exist regulated by repressor proteins that bind to specific sequences in their iii´ untranslated regions. These regulatory proteins may also direct mRNAs to specific regions of eggs or embryos, allowing localized synthesis of the encoded proteins during embryonic development.
![]()
Box
Key Experiment: Catalytic Function of Ribosomal RNA.
![]()
Box
Molecular Medicine: Antibiotics and Protein Synthesis.
hillyerinctiary1996.blogspot.com
Source: https://www.ncbi.nlm.nih.gov/books/NBK9849/#:~:text=All%20mRNAs%20are%20read%20in,a%20nearly%20universal%20genetic%20code.
Belum ada Komentar untuk "Which Way Is Rna Read During Translation"
Posting Komentar